ОТ ФЕНОМЕНА ЧЕЛОВЕКА К ЧЕЛОВЕЧЕСКОЙ СУЩНОСТИ
I. ПРЕДЖИЗНЬ
II. ЖИЗНЬ
III. МЫСЛЬ
IV. СВЕРХЖИЗНЬ
ЭКСПАНСИЯ ЖИЗНИ
Когда физик хочет исследовать распространение волны, он начинает вычислять пульсацию одной частицы. Затем, сведя вибрирующую среду к ее основным характеристикам, как и главным направлениям упругости, он обобщает найденные для данного элемента результаты в соответствии с этой средой. Таким путем он получает максимально приближенные основные очертания совокупного движения, которое он стремился определить.
Примерно так же, только своими собственными средствами, вынужден действовать и биолог, вставший перед задачей описать подъем жизни. Нельзя установить порядок в этом огромном и сложном феномене, не проанализировав предварительно способы, изобретенные жизнью для продвижения вперед, в каждом из ее элементов по отдельности. И невозможно выявить общий вид этих многих суммированных индивидуальных прогрессов, не отобрав в их результирующей наиболее выразительные и ясные черты.
Упрощенное, но структурное представление о развивающейся земной жизни. Видение, истинность которого выступает в чистом и неотразимом эффекте однородности и цельности. Без второстепенных деталей и дискуссий. Еще и еще раз перспектива, которую надо видеть и признать или же не видеть совсем. Вот что я предполагаю развить в последующих параграфах.
Суть того, что я хочу сказать, содержится и определяется в трех главных разделах:
1. Элементарные формы движения жизни.
2. Спонтанное разветвление живой массы.
3. Древо жизни.
Все это сначала рассматривается с внешней стороны, с поверхности. И только в следующей главе мы попытаемся проникнуть во внутреннюю сторону вещей.
1. ЭЛЕМЕНТАРНЫЕ ФОРМЫ ДВИЖЕНИЯ ЖИЗНИ
А) Самовоспроизведение
В основе всего процесса образования вокруг Земли оболочки биосферы лежит типично жизненный механизм самовоспроизведения. Всякая клетка в определенный момент делится (путем «бинарного деления», или «кариокинеза») и порождает новую, схожую с ней клетку. Был только один центр, теперь их стало два. Все последующие движения жизни вытекают из этого элементарного и мощного феномена.
Само по себе деление клетки, видимо, вызывается просто необходимостью для живой частицы избавиться от своей молекулярной неустойчивости и от структурных трудностей, связанных с продолжением своего возрастания. Омоложение и облегчение. Ограниченные группировки атомов - микромолекулы имеют почти бесконечную долговечность (но зато и почти бесконечную неизменность). Клетка, постоянно ассимилируя, должна для продолжения существования делиться надвое. Поэтому самовоспроизведение вначале оказывается простым способом, изобретенным природой для обеспечения постоянства неустойчивого в случае крупных молекулярных построений.
Но, как это всегда бывает в мире, то, что вначале явилось лишь счастливым случаем или способом выжить, было немедленно преобразовано и использовано как орудие прогресса и завоевания. Вначале жизнь воспроизводилась, видимо, лишь для самозащиты. Но это оказалось прелюдией к ее захватам.
Б) Размножение
Ибо, будучи однажды введенным в ткань универсума, принцип удвоения живых частиц не знает иных границ, кроме количества материи, подверженной его действию. Подсчитано, что через несколько поколений одна-единственная инфузория путем простого деления самой себя и своего потомства покрыла бы всю Землю. Никакой самый большой объем не выдерживает воздействия геометрической прогрессии. И это не простая экстраполяция рассудка. Благодаря одной только способности к постоянному удвоению, которой ничто не может воспрепятствовать, жизнь обладает столь же неодолимой силой экспансии, как и тело, которое расширяется или испаряется. Но, если в случае так называемой инертной материи увеличение объема вскоре достигает точки равновесия, то в случае живого вещества не обнаруживается никакого снятия напряжения. Чем больше распространяется явление клеточного деления, тем активнее оно становится. Раз начавшуюся игру раздвоения ничто изнутри не остановит, ибо этот строяще-пожирающий огонь спонтанен. А следовательно, и вовне ничто не является достаточно крупным, чтобы затушить его путем насыщения.
В) Обновление
Но это лишь только первый результат и количественная сторона происходящей операции. Самовоспроизведение удваивает материнскую клетку. И так, путем механизма, обратного химическому разложению, происходит умножение без раздробления. Но, кроме того, одновременно преобразуется то, что должно было быть лишь только продолжено. Замкнутый в себе, живой элемент более или менее быстро приходит в состояние неподвижности. Он застревает и застывает в своей эволюции. В момент самовоспроизведения и под его действием он вновь обретает способность внутренне перестраиваться и, следовательно, принимать новый облик и новую направленность. Умножение по форме, как и по числу. Начальная волна жизни, исходящая от каждого индивида, распространяется не как монотонный круг из других, совершенно сходных с ним индивидов. Она преломляется и переливается бесконечной гаммой оттенков. Центр неодолимого умножения, живое существо тем самым становится не менее неодолимым очагом разнообразия.
Г) Спаривание
И тогда-то, как будто затем, чтобы расширить брешь, пробитую ее первым потоком в стене неорганизованного, жизнь открыла чудесный способ спаривания. Потребовался бы целый труд, чтобы определить и восхититься тем, как возрастает и утончается путем эволюции от клетки до человека разделение полов. На начальном этапе, где мы его сейчас рассматриваем, это явление выступает в основном как средство ускорить и усилить двойное действие умножения и разнообразия, достигнутое сначала бесполым размножением, как оно еще функционирует у многих низших организмов, вплоть до каждой клетки нашего тела. Путем первого спаривания двух элементов (как бы мало они ни были еще дифференцированы на мужской и женский) открылась дверь к тем способам размножения, при которых один индивид может распылиться в мириады зародышей. И одновременно началась бесконечная игра - комбинирование «признаков», анализ которого тщательно проводится современной генетикой. Вместо того чтобы просто расходиться из каждого делящегося центра, лучи жизни начали с этого времени соединяться своими концами, обмениваясь своими богатствами и разнообразя их. Не удивляясь изобретению огня, хлеба или письменности, мы не думаем удивляться и этому изумительному изобретению. И, однако, потребовалось столько случайностей и столько попыток - и, следовательно, столько времени, - прежде чем созрело это фундаментальное открытие, из которого вышли мы сами! А сколько еще понадобилось времени, пока оно не нашло свое естественное дополнение и завершение в не менее революционном новшестве ассоциации!
Д) Ассоциация
При первом анализе, еще не затрагивая более глубоких факторов, группирование живых частиц в сложные организмы представляется почти неизбежным следствием их размножения. Клетки стремятся к агломерации потому, что они прижимаются друг к другу или даже возникают группами. Но из этой чисто механической целесообразности или необходимости сближения в конечном итоге был порожден и выделился определенный метод биологического совершенствования.
Сохранились и существуют на наших глазах в природе, кажется, все стадии этого еще не законченного движения к единению или синтезу беспрерывно увеличивающихся продуктов размножения живого. В самом низу - простая агрегация, как это имеет место у бактерий или низших грибов. На следующей, более высокой ступени - спаянная колония с более четко специализированными, но еще совсем не централизованными элементами - таковы высшие растения, мшанки или полипы.
Еще выше - многоклеточное - настоящая клетка клеток, у которой путем поразительного коренного преобразования над организованной группой живых частиц - как бы из-за чрезмерной скученности - устанавливается самостоятельный центр. И, наконец, еще выше, у нынешней границы нашего опыта и экспериментов жизни - общества, эти загадочные ассоциации свободных многоклеточных, в недрах которых делаются более или менее удачные попытки создания путем «мегасинтеза» сверхсложных единств.
Последняя часть этой книги будет специально посвящена исследованию этой последней и высшей формы группирования, в которой стремление материи к организации достигает, вероятно, кульминационного пункта в разумно организованном обществе (Social reflechi). Ограничимся здесь замечанием, что ассоциация у одушевленных существ на всех ее ступенях - не спорадическое или случайное явление. Напротив, она представляет собой один из самых универсальных, постоянных и, следовательно, самых знаменательных механизмов, используемых жизнью для своей экспансии. Из ее преимуществ непосредственно очевидны два. Прежде всего благодаря ассоциации живое вещество организуется в массы, достаточно крупные, чтобы вырваться из множества внешних зависимостей (капиллярное сцепление, осмотическое давление, химическое изменение среды и т. д.), которые парализуют микроскопическое существо. В биологии, как и в навигации, физически требуется определенный размер, чтобы некоторые движения стали возможными... И также благодаря ассоциации (опять-таки вследствие получающегося увеличения в объеме) организм находит внутри себя достаточно места, чтобы разместить многочисленные детали механизма, возникшие путем постепенного прибавления из его дифференциации.
Е) Направленное прибавление
Самовоспроизведение, спаривание, ассоциация... Сколько бы ни продолжались эти различные формы движения клетки, сами по себе они предопределяют лишь развертывание организмов на поверхности. Если бы жизнь свелась только к ним. то она распространялась бы и разнообразилась лишь в одной плоскости. Она походила бы на самолет, который бежит по дорожке, будучи не в силах «оторваться». Она не смогла бы подняться.
Здесь-то и вступает в силу, играя роль вертикальной слагающей, явление прибавления.
Конечно, в ходе биологической эволюции имеется немало примеров превращений, идущих по горизонтали, путем чистого перекрещивания признаков. Таковы так называемые менделевские мутации. Однако чаще обновления, ставшие возможными при каждом размножении, дают нечто лучшее, чем взаимную замену, они прибавляются одни к другим, и их сумма растет в определенном направлении. Предрасположения, которые усиливаются, или органы, которые пригоняются или накладываются друг на друга. Здесь - возрастающее разнообразие, там - увеличивающаяся специализация членов, образующих единый генеалогический ряд. Иными словами, появление линии развития (lignee) как естественной единицы, отличной от индивида. Этому закону направленного усложнения, в ходе которого вызревает и сам процесс, породивший, исходя из микромолекул, а затем мегамолекул, первые клетки, биология дала название ортогенеза1.
Ортогенез - динамическая и единственно полная форма наследственности. Какая реальность и какие пружины космического масштаба скрываются за этим словом? Мы постепенно это выясним. Но уже на этой стадии нашего исследования ясно виден первый пункт. Из-за характерной для него способности к прибавлению живое вещество (в противовес материи физиков) «страдает» сложностью и неустойчивостью. Оно падает, или, вернее, поднимается, ко все менее вероятным формам.
Без ортогенеза было бы лишь распространение вширь; с ортогенезом обязательно имеется какой-то подъем жизни.
Одно из следствий. Способы действия жизни
Теперь приостановимся на один момент. И прежде чем исследовать, как проявляются в применении к жизни в целом различные законы, установленные выше для отдельной частицы, попытаемся выяснить, каковы в силу этих самых законов элемента общие способы действия или общие положения, которые на всех уровнях и при всех обстоятельствах характеризуют движущуюся жизнь.
Эти положения или способы действия могут быть сведены к трем: изобилие, изобретательность и (с нашей индивидуальной точки зрения) безразличие.
а) Изобилие. Оно возникает из безграничного процесса размножения. Жизнь действует путем эффекта масс, посредством множеств, брошенных, казалось бы, в беспорядке вперед. Миллиарды зародышей и миллионы взрослых особей теснят друг друга, отталкивают, пожирают - кто займет больше места и лучшие места.
Видимое расточительство и ожесточенность, тайна и скандал и вместе с тем, надо признать, биологическая эффективность борьбы за существование. В ходе безжалостной игры, которая сталкивает и втискивает друг в друга массы живого вещества, находящиеся в процессе безудержного расширения, индивид наверняка доходит до пределов своих возможностей и своего усилия. Выживание наиболее приспособленного, естественный отбор - это не пустые слова, если не вкладывать в них ни конечный идеал, ни окончательное объяснение.
Но в этом явлении главную роль играет, по-видимому, не индивид- Более глубоким фактором борьбы за существование является битва шансов, а не ряд одиночных боев. Размножаясь в бесчисленности, жизнь делает себя неуязвимой для наносимых ей ударов. Она увеличивает свои шансы выжить. И одновременно она умножает свои шансы на продвижение вперед.
И вот здесь-то продолжается, выступая на уровне одушевленных частиц, важнейшая техника пробного нащупывания - это специфическое и неотразимое оружие всякого расширяющегося множества. В пробном нащупывании весьма любопытно сочетаются слепая фантазия больших чисел и определенная целенаправленность. Пробное нащупывание - это не просто случай, с которым его хотели смешать, но направленный случай. Все заполнить, чтобы все испробовать. Все испробовать, чтобы все найти, И то, что природа, если можно так выразиться, ищет путем создания изобилия, не есть ли это, по существу, возможность применить этот прием, становящийся все более громоздким и дорогостоящим по мере того, как он больше охватывает.
б) Изобретательность. Это - необходимое условие, или. точнее, конструктивная сторона прибавления.
Чтобы объединять свойства в устойчивые и цельные комбинации, жизнь должна проявлять исключительную ловкость. Ей надо выдумывать и комбинировать детали механизма в минимуме пространства. Подобно инженеру, она должна собирать механизмы гибкие и простые. Но из этого вытекает, что структура организмов (чем более они высокоорганизованы!) обладает свойством, о котором никогда нельзя забывать.
Что собирается, то и разбирается.
На первой стадии своих открытий биология была удивлена и заворожена тем фактом, что живые существа, какой бы совершенной или даже чем более совершенной ни была их спонтанность, под ее пальцами всегда поддавались разложению в бесконечную цепь замкнутых механизмов. Отсюда был сделан вывод о всеобщности материализма. Но это значило забыть о существенном различии между естественным целым и продуктами его анализа.
Верно, что по строению любой организм всегда и необходимо разбирается на составные части. Но из этого отнюдь не следует ни что само суммирование этих частей происходит автоматически, ни что из их суммы не возникает чего-то специфически нового.
То, что «свободное», даже у человека, оказывается целиком разложимым на детерминированные части, не опровергает того, что в основе мира (как мы здесь утверждаем) лежит свобода. Это просто результат и триумф изобретательности жизни.
в) Безразличие к индивидам. Сколько раз искусство, поэзия и даже философия рисовали природу в виде женщины с завязанными глазами, топчущей бесчисленное множество раздавленных жизней... Первый след этой кажущейся жестокости проявляется в изобилии. Как саранча, о которой писал Толстой, жизнь проходит по мосту из нагроможденных трупов. И это - прямое следствие размножения. Но в том же «бесчеловечном» направлении по-своему действуют и ортогенез и ассоциация.
Путем явления ассоциации живая частица как бы отрывается от самой себя. Оказываясь в более обширной, чем сама, совокупности, она частично попадает в рабство этой совокупности. Она уже не принадлежит самой себе.
И если включение в органическое или социальное растягивает частицу в пространстве, то приобщение к линии потомков не менее неумолимо растягивает ее во времени. Ортогенез - волочильня для индивида. Из центра он становится посредником, звеном. Он больше не существует как таковой - он передает. Жизнь более реальна, чем жизни, было уже сказано...
Здесь - гибель во множестве. Там - четвертование в коллективе. А еще в третьем направлении - растягивание на дыбе становления. Драматическое и вечное противоречие между возникшим из множества элементом и множеством, постоянно возникающим из элемента в ходе эволюции.
По мере того как общее движение жизни упорядочивается, этот конфликт, несмотря на его периодические вспышки, идет к своему разрешению. Однако его безжалостное действие распознается все же до самого конца. И только на стадии духа, когда эта антиномия доходит до пароксизма, воспринимаемого чувством, она проясняется, и тогда безразличие мира к своим элементам превращается в огромную заботу - в сфере личности.
Но мы до этого еще не дошли.
Нащупывающее изобилие, созидательная изобретательность, безразличие ко всему, что не является будущностью и совокупностью. Таковы три тенденции, под знаком которых в силу своих элементарных механизмов жизнь поднимается все выше и выше. Но есть еще и четвертая, объемлющая первые три, - тенденция всеохватывающего единства.
Эту последнюю мы уже встречали в первородной материи, затем, на молодой Земле, потом при «вылупливании» первых клеток. Здесь она проявляется еще раз и со все большей очевидностью. Как бы широко и многообразно ни разрасталась живая материя, распространение ее ростков всегда происходит солидарно. Внешне они постоянно прилаживаются, приспосабливаются друг к другу. Внутренне они глубоко уравновешиваются. Взятое в целом, живое вещество, расползшееся по Земле, с первых же стадий своей эволюции вырисовывает контуры одного гигантского организма.
В конце каждого из этапов, которые ведут нас к человеку, я, как припев, все повторяю одно и то же. Потому что, стоит только забыть об этом, все станет непонятным.
Чтобы увидеть жизнь за множеством индивидуальных существований и их соперничеством, ни в коем случае нельзя упускать из виду единство биосферы. Вначале еще расплывчатое единство. Скорее единство происхождения, общих рамок, рассеянного порыва, чем единство упорядоченной группировки. Но по мере подъема жизни оно делается все более определенным, свертывается на себя и, наконец, на наших глазах обретает центр.
2. РАЗВЕТВЛЕНИЯ ЖИВОЙ МАССЫ
Посмотрим теперь, как выглядят на всем пространстве живой земли те формы движения, которые мы проанализировали у отдельных клеток и их отдельных группировок. Можно было бы представить себе, что, взятое в таких размерах, все это множество перепутается и породит лишь отчаянный беспорядок. Или, наоборот, что их сумма, придя в гармонию, даст своего рода непрерывную волну, как та, что возникает в спокойной воде от брошенного камня. В действительности же не происходит ни то, ни другое. В той форме, в какой он и сейчас предстает перед нашим взором, фронт восходящей жизни не является ни беспорядочным, ни непрерывным. Он выступает как совокупность фрагментов, одновременно расходящихся и идущих по ступеням - классы, отряды, семейства, роды, виды. Это та самая последовательность групп, разнообразие, порядковую величину и сцепление которых пытается выразить в своей номенклатуре современная систематика.
Рассматриваемая в целом, жизнь при продвижении вперед расчленяется. В своей экспансии она самопроизвольно распадается на широкие естественные подразделения, образующие иерархию. Она разветвляется. Таков специфический феномен, столь же существенный для больших живых масс, как для клеток «кариокинез». Пришло время им заняться.
Имеется целый ряд факторов, в той или иной степени участвующих в формировании и укреплении ветвистой системы жизни. Я их сведу к следующим трем:
а) сцепления роста, порождающие «филы»;
б) распускания (или разъединения) зрелости, периодически производящие «мутовки»;
в) действие отдаленности, с виду устраняющее «черешки».
А. Сцепления роста
Вернемся к живому элементу, находящемуся в процессе самовоспроизведения и размножения. Мы видели, что из этого элемента, как центра, в силу ортогенеза во все стороны расходятся различные линии потомства, каждая из которых узнается по усилению некоторых признаков. По своему строению эти линии расходятся и стремятся отдалиться друг от друга. Однако еще ничто не указывает на то, что путем встречи с потомствами, происшедшими из соседних элементов, они не смещаются настолько, что вместе образуют непроницаемое сплетение.
Под «сцеплением роста» я подразумеваю тот новый и неожиданный факт, что там, где можно было бы ожидать образования игрой случая сложного переплетения, происходит как раз рассеивание простого типа. Разлитая на земле вода сразу же растекается ручейками, затем вполне оформленными ручьями. Подобно этому, под влиянием различных причин (врожденный параллелизм ортогенеза элементов, взаимное тяготение и приспособление друг к другу потомственных линий, избирательное действие среды...) волокна живой массы, разнообразясь, стремятся сблизиться, сгруппироваться, сплестись по нескольким доминирующим направлениям. Вначале эта концентрация форм вокруг нескольких главных осей выступает еще неотчетливо и расплывчато - как простое возрастание на некоторых участках числа и густоты линий. А затем мало-помалу она усиливается. Вырисовываются настоящие прожилки, но они пока не нарушают формы листа, в середине которого появились. В это время волокна частично еще ускользают от сети. которая стремится их захватить. От прожилки к прожилке они еще могут соединиться, сомкнуться и переплестись. Группировка, скажет зоолог, находится еще на стадии расы. И вот тогда-то одновременно происходит, в зависимости от того, как смотреть, окончательное сцепление или разъединение. Достигнув определенной степени взаимосвязи, линии обосабливаются в замкнутый пучок, отныне непроницаемый для соседних пучков. Их ассоциация впредь будет эволюционировать сама по себе, как нечто самостоятельное. Вид индивидуализировался. Возникла фила.
Фила. Живой пучок. Линия линий. Многие еще не желают видеть или считать реальным это звено развивающейся жизни. Но они не умеют приспособить свое зрение, не хотят как следует смотреть.
Прежде всего фила - это коллективная реальность. Поэтому для того, чтобы четко ее различить, надо стать достаточно высоко и далеко. На слишком близком расстоянии она размельчается на нечто неопределенное и беспорядочное. За деревьями не виден лес.
Далее, фила полиморфна и эластична. Подобно молекуле, которая может иметь любые размеры и любые степени сложности. фила может быть мелкой, как вид, или крупной, как тип. Бывают простые филы и филы фил. Филетическое единство более структурное, чем количественное. Поэтому надо уметь узнавать его при любом размере.
Наконец, фила по своей сущности - динамическая реальность. Она ясно выступает лишь при определенной глубине длительности, то есть в движении. Будучи остановленной во времени, она теряет свое лицо и как бы свою душу. Жест на фотоснимке мертв.
При рассмотрении без этих оговорок фила кажется лишь еще одной искусственной сущностью, вырезанной в целях классификации из живого континуума. При наблюдении с необходимым увеличением и в соответствующем свете она, напротив, раскрывается как совершенно определенная структурная реальность.
«Первоначальный угол расхождения», то есть то особое направление, в котором она группируется и развивается, отделяясь от соседних форм, - вот что, во-первых, определяет филу.
Во-вторых, ее определяет ее «первоначальное сечение».
По этому последнему пункту (который уже затрагивался в связи с первыми клетками и будет иметь большое значение, когда речь пойдет о человеке) нам еще почти ничего не известно. Но ясно по крайней мере одно. Как капля воды физически способна сконденсироваться, лишь начиная с определенного объема, или как химическое превращение может начаться лишь при определенном количестве участвующего вещества, так и фила может биологически сформироваться лишь тогда, когда она с самого начала группирует в себе достаточно большое число потенций и притом достаточно разнообразных. Без достаточной густоты и первоначального изобилия (как и без достаточного отклонения при старте) никогда, мы теперь это знаем, новая ветвь не сумеет индивидуализироваться. Это - совершенно четкое правило. Но как оно конкретно действует и выражается? Диффузная сегрегация одной массы внутри другой? Возбуждение, распространяющееся вокруг сугубо ограниченной мутационной зоны? В какой форме представить себе на поверхности возникновение какого-либо вида? Мы еще колеблемся. И вопрос, быть может, влечет разные ответы. Но суметь ясно поставить проблему не значит ли почти разрешить ее?
Наконец, завершает определение филы и, кроме того, недвусмысленно помещает ее в категорию естественных единиц мира «ее способность к самостоятельному развитию и специфический закон этого развития». Хотя и по-своему, но фила действительно ведет себя как живое существо - она растет и распускается, как цветок.
Б. Распускания зрелости
В силу аналогии, которая, как мы далее откроем, основывается на глубокой природной связи, развитие филы идет удивительно параллельно последовательным стадиям, через которые проходит человеческое изобретение.
Мы хорошо знаем эти стадии, так как постоянно их наблюдали на протяжении столетия вокруг нас. Вначале идея оформляется приблизительно в теории или в механизме. Затем идет период быстрых модификаций - беспрерывных поправок и переделок до более или менее окончательной отработки. Достигнув этой стадии, новое создание вступает в фазу распространения и равновесия. Качественно оно изменяется лишь в некоторых второстепенных деталях - оно «достигло потолка». Зато оно распространяется и полностью закрепляется количественно. Такова история всех современных изобретений - от велосипеда до самолета, от фотографии до кино и радиовещания.
Схожей вырисовывается в глазах естествоиспытателя кривая развития ветвей жизни. Вначале фила - это «открытие» путем пробного нащупывания нового органического типа, обладающего жизнеспособностью и определенными преимуществами. Но этот тип не сразу достигает наиболее экономичной и наилучше приспособленной формы. В течение более или менее длительного времени он употребляет всю свою силу, так сказать, на испробование самого себя. Пробы следуют друг за другом, однако окончательно еще не закрепляются. Но вот наконец близится совершенство. С этого момента темп изменений замедляется, и новое создание, достигнув пределов того, что оно может дать, вступает в фазу завоевания.
Более сильная, чем ее менее усовершенствованные соседи, вновь возникшая группа распространяется и одновременно закрепляется. Она размножается, но без дальнейшего изменения. Она одновременно достигает максимума величины и максимума устойчивости.
Распускание филы путем простого расширения или путем простого утолщения ее первоначального стебля. За исключением случая ветви, достигшей пределов своей эволюционной мощи, этот элементарный случай никогда строго не реализуется. Как бы успешно ни разрешила новая форма проблемы, поставленные существованием, это решение допускает некоторое число вариантов, каждый из которых имеет свои преимущества и которые не имеют никакого основания и никакой возможности взаимно устранить друг друга. Этим объясняется тот факт, что по мере своего утолщения фила стремится разъединиться на вторичные филы, каждая из которых представляет собой один вариант, или обертон, основного типа. Она как бы расщепляется вдоль своего фронта расширения. Одновременно с количественным распространением она качественно подразделяется. Это возобновляется разъединение. Новые подразделения то кажутся выражением лишь поверхностного разнообразия, результатом случая или буйной фантазии. То, напротив, они представляют собой точные приспособления основного типа к конкретным нуждам или конкретным зонам распространения. Так появляются лучи («радиации»), столь явственно обозначенные, как мы скоро увидим, у позвоночных. Как и следовало ожидать, внутри каждого луча механизм стремится повториться, хотя и в ослабленном виде. Лучи, таким образом, в свою очередь проявляют признаки веерообразного членения. Теоретически процесс бесконечен. В действительности же опыт показывает, что это явление вскоре затухает. Образование вееров довольно быстро прекращается, и дальнейшее расширение ветвей на концах происходит уже без заметного их разделения.
В своем самом общем виде распустившаяся фила - это в конечном итоге мутовка закрепившихся форм.
Но тут, придавая последний штрих всему феномену, в центре каждой части мутовки обнаруживается глубокая склонность к социализации. О социализации следует повторить то, что выше говорилось вообще о жизненной силе ассоциации. Поскольку в природе оформленные группировки индивидов, или организованные и дифференцированные сообщества, относительно редки (термиты, перепончатокрылые, люди...), мы склонны видеть в них какую-то исключительную черту эволюции. В противовес этому первому впечатлению при более внимательном наблюдении вскоре выясняется, что они выражают собой один из наиболее существенных законов организованной материи. Последний метод, которым живая группа, делаясь более сплоченной, усиливает свою сопротивляемость уничтожению и свою способность к завоеванию? Полезный способ, изобретенный ею особенно для увеличения своего внутреннего богатства путем совместного объединения ресурсов?.. Какова бы ни была его глубокая причина - факт налицо. Когда в конце каждого побега мутовки элементы филы обретают свою окончательную форму, они столь же определенно стремятся к сближению и социализации, как атомы твердого тела стремятся к кристаллизации.
Когда же достигнутый таким образом прогресс реализуется в укреплении и индивидуализации концов ее веера, то можно сказать, что фила достигла своей полной зрелости. С этого момента она будет существовать до тех пор, пока из-за внутреннего ослабления или внешнего соперничества не поредеет и не устранится окончательно. Тогда, за исключением случайно выживших нескольких навсегда застывших в своем развитии линий, ее история заканчивается, если только путем явления самооплодотворения она не сумеет в том или ином месте выпустить новую почку.
Чтобы понять механизм этой способности оживления, нужно возвратиться к идее, или символу, нащупывания. Образование мутовки, сказали мы, объясняется прежде всего необходимостью для филы множиться, чтобы соответствовать различным потребностям или возможностям. Но благодаря тому, что число лучей увеличивается и каждый разворачивающийся новый луч увеличивает, сверх того, число индивидов, то множатся также и «пробы», «опыты». Веер в конце филы - это лес исследующих щупальцев. Стоит лишь одному из этих щупальцев случайно найти щель или формулу, открывающую доступ в новое отделение жизни, - и тогда, вместо того чтобы закрепляться или достигать потолка в монотонных различиях, ветвь в этой точке вновь обретает свою подвижность. Она вступает в состояние мутации. Открывается путь для новой пульсации жизни, которая вскоре под влиянием комбинированных сил сцепления и разъединения в свою очередь делится на мутовки. Возникает новая фила, которая растет и распускается над ответвлением, на котором возникла, не обязательно заглушая и истощая его. Быть может, пока на ней самой не вырастет третья ветвь, затем четвертая, если, конечно, взято правильное направление и если это позволяет общее равновесие биосферы.
В. Действие отдаленности
Таким образом, самим ритмом своего развития каждая линия жизни попеременно то сужается, то расширяется. Цепь «сгущений» и «разряжений», ряд тонких черешков и широких листьев - таков ее облик.
Но эта схема дает лишь теоретическое представление о происходящем. Увидеть ее таковой мог бы только земной наблюдатель, одновременно присутствующий на протяжении всей длительности. Но такой наблюдатель - лишь вообразимая нелепость. В действительности подъем жизни может быть нами обнаружен, лишь исходя из очень краткого отрезка, т. е. сквозь огромную толщу истекшего времени. Значит, то, что дано нам в опыте и что, следовательно, составляет «феномен», - это не эволюционное движение само по себе, а эволюционное движение с добавлением искажений, вызванных действием отдаленности. Но в чем выражаются эти искажения? Попросту в усилении (быстро возрастающем с расстоянием) веерообразности структуры, возникшей из филетического лучеиспускания жизни. Это происходит, впрочем, двумя различными способами - во-первых, путем еще большего увеличения видимого рассеивания фил и, во-вторых, путем видимого устранения их черешков.
Еще большее увеличение видимого рассеивания фил. Это первое действие перспективы, видимое всем, связано с постарением и «прореживанием» ветвей жизни в результате возраста. В настоящее время в природе сохранилось лишь весьма незначительное число организмов, последовательно выросших на стволе жизни. И как бы ни старалась палеонтология, много угасших форм останутся для нас навсегда неизвестными. В результате этого истребления в кроне растительных и животных форм все время образуются просветы. Эти просветы становятся все шире по мере того, как мы спускаемся к истокам. Обламываются сухие ветки. Опадают листья. Все это исчезающие посредующие морфологические звенья, отсутствие которых столь часто придает выжившим линиям вид голых и одиноких стеблей. Одна и та же длительность одной рукой умножает свои творения впереди, а другой рукой не менее уверенно разрежает их позади. Таким образом, она их все более разделяет, все более изолирует, на наш взгляд, тогда как с помощью другого, более тонкого приема она создает у нас иллюзию того, что они плавают, как облака, без опоры, над бездной прошедших веков.
Устранение черешков. С героических времен Ламарка и Дарвина излюбленная тактика в борьбе против трансформистов всегда состояла в напоминании им о том, что они бессильны доказать возникновение вида на материальных следахi. «Несомненно, - говорят им, - вы показываете последовательность различных форм в прошлом и даже, мы с этим согласны, в некоторых пределах превращение этих форм друг в друга. Но как бы примитивны они ни были, ваше первое млекопитающее - это все-таки уже млекопитающее; ваша первая лошадь (Equide) - уже лошадь (cheval), и так далее. Может быть, имеется эволюция внутри типа. Но нет появления типа путем эволюции». Так и поныне продолжают твердить все более редкие сторонники неизменности.
Как будет видно, независимо от аргументов, вытекающих из постоянно накапливающихся очевидных фактов палеонтологии, имеется более радикальный ответ на это возражение (или скорее. категорическое отклонение его) - это отрицание его предпосылки. В сущности, антитрансформисты требуют, чтобы им показали «черешок» филы. Но это столь же неразумное, сколь и бесполезное требование, ибо для его удовлетворения потребовалось бы изменить сам строй мира и условия нашего восприятия.
По природе нет ничего более деликатного и мимолетного, чем начало. До тех пор пока зоологическая группа молода, ее признаки остаются неопределенными. Ее сооружение хрупко. Ее размеры малы. Ее составляют относительно немного индивидов, и они быстро изменяются. Как в пространстве, так и в длительности черешок (или. что то же самое, почка) живой ветви обладает минимумом дифференциации, экспансии и сопротивляемости. Как же действует время на эту слабую зону?
Неизбежно уничтожая то, что от нее остается.
Эту раздражающую, но существенную непрочность исходного материала должны почувствовать все, кто занимается историей!
В любой области, когда вокруг нас начинает чуть пробиваться что-то действительно новое, мы его не замечаем по той простой причине, что нам надо было бы видеть его расцвет в будущем, чтобы заметить его в самом начале. А когда та же самая вещь выросла и мы оборачиваемся назад, чтобы найти ее зародыш и первые наброски, то тогда в свою очередь скрываются эти первые стадии, уничтоженные или забытые. Где столь близкие, однако, к нам древние греки или древние римляне? Где первые прялки, первые колесницы и первые очаги? Где (уже) первые модели автомобилей, самолетов, киноаппаратов?.. Везде - в биологии, культуре, лингвистике. - как резинка в руках художника, время стирает каждую бледную линию в рисунке жизни. Путем механизма, детали которого в каждом отдельном случае кажутся устранимыми и случайными, но универсальность которого доказывает, что он отражает фундаментальное условие нашего знания, зародыши, черешки, первоначальные фазы роста, каковы бы они ни были, исчезают с наших глаз позади. Кроме закрепившихся максимумов, кроме упрочившихся завершений, ничто (ни в виде «свидетелей», ни даже в виде следов) не остается от того, что было до нас. Иначе говоря, только конечные лучи вееров доходят до нынешнего времени или как живые представители, или в ископаемом виде.
Поэтому нет ничего удивительного в том, что ретроспективно вещи кажутся нам появившимися в готовом виде2.
Автоматически, путем избирательного поглощения веков развитие стремится исчезнуть с нашего горизонта и превратиться во всей области явлений в прерывную последовательность плоскостей и постоянств3 ii.
Так, под разрушительным действием прошлого, сочетающимся с созидательным действием роста, окончательно вырисовываются и выделяются для науки разветвления древа жизни.
Попробуем рассмотреть это древо в его конкретной реальности и измерить его.
3. ДРЕВО ЖИЗНИ
А. Основные черты
а) Одна из количественных единиц эволюции - пласт млекопитающих
Из предшествующих замечаний непосредственно вытекает, что для четкого представления древа жизни необходимо «натренировать взгляд» на той части его кроны, на которой лишь умеренно отразилось разрушающее действие времени. Ни слишком близко, чтобы листья не мешали видеть; ни слишком далеко, чтобы ветви были еще достаточно густыми.
Где найти в нынешней природе этот привилегированный участок? Безусловно, в большой семье млекопитающих.
Мы положительно знаем из геологии (достаточно обратить внимание на ее внутреннюю структуру), что хотя человечество еще «незрелая» группа, в целом млекопитающие - группа взрослая и в то же время свежая. Они полностью развились лишь в третичном периоде, и среди них до сих пор сохранилось значительное количество его самых деликатных остатков. Вот почему эта группа с самого начала была и остается излюбленным объектом при возникновении и развитии трансформистских идей.
Рассмотрим эту группу в ее основных чертах (рис. 1). Однако для начала ограничим поле наших исследований самой молодой и прогрессивной частью этой группы: плацентарными млекопитающими4.
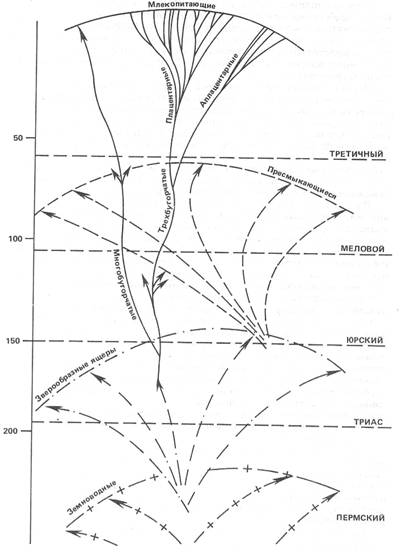
Рис. 1. Схема, символизирующая развитие слоями четвероногих. Цифры слева обозначают миллионы лет. Детальное объяснение в тексте.
С эволюционной (можно даже сказать, с «физиологической») точки зрения плацентарные млекопитающие, взятые в целом, составляют то, что я условно здесь называю биот. Под этим я подразумеваю мутовочную группировку, элементы которой не только родственны по рождению, но и, кроме того, поддерживают и взаимно дополняют друг друга в борьбе за существование и распространение.
Чтобы понять этот важный пункт, выясненный главным образом американской школой палеонтологииiii, достаточно рассмотреть в соответствующем свете распределение наиболее знакомых каждому из нас животных форм. Здесь - травоядные и грызуны, извлекающие свою пищу непосредственно из растительного ответвления жизни; там - насекомоядные, паразитирующие подобным же образом на «членистоногих». Здесь - плотоядные, существующие за счет тех и других, а там - всеядные, питающиеся за всеми столами сразу. Таковы четыре главные радиации, в основном совпадающие с общепринятым делением фил.
Рассмотрим теперь эти четыре луча или сектора один за другим, раздельно. Каждый из них совершенно свободно подразделяется, расщепляется на меньшие подразделения. Возьмем, например, ту ветвь, которая сейчас представляется самой густой, - травоядных. В зависимости от двух различных способов, избранных для превращения конечностей в бегающие лапы (путем исключительного развития двух пальцев или одного среднего пальца), появляются два крупных семейства - парнокопытные и непарнокопытные, каждое из которых образуется из пучка отдельных больших линий. Здесь у непарнокопытных - серая толпа тапирообразных, короткая, но удивительная ветвь титанотериев, халикотерии с землероющими когтями, которых человек, вероятно, еще застал, племя носороговых, безрогих или с рогом, и, наконец, однокопытные лошади с параллельной им совершенно независимой филой в Южной Америке. Там, у парнокопытных - свиньи, верблюды, олени и антилопы, не говоря уже о других стеблях, менее жизнеустойчивых, но столь же индивидуализированных и интересных с точки зрения палеонтологии. А ведь есть еще обширная и мощная группа хоботных! Согласно правилу «устранения черешков», начало каждого из этих подразделений тонет в тумане прошлого.
Но раз уж они появились, то мы можем проследить, у всех вместе и у каждого в отдельности главные фазы их географического распространения; их последовательное деление, почти до бесконечности, на субмутовки; наконец, чрезмерное усиление путем ортогенеза некоторых особенностей костей, зубов или черепа, что обычно превращает животных в уродов или делает их недолговечными.
Но и это еще не все. Поверх этого обилия родов и видов, образовавшихся из четырех основных радиации, мы различаем другую сеть, соответствующую сделанным там и сям попыткам покинуть наземную жизнь и оккупировать воздух, воду и даже глубины земли. Наряду с формами, приспособленными для бега, появляются лазающие и летающие, плавающие и землероющие. Одни (киты и сирены), очевидно, с удивительной быстротой произошли от хищных и травоядных. Другие (рукокрылые, кроты и земляные крысы) - главным образом от наиболее древних элементов плацентарной группы - насекомоядных и грызунов, существующих с конца вторичного периода.
Рассматривая этот столь хорошо сбалансированный функциональный ансамбль сам по себе, нельзя не признать, что он представляет собой органическую естественную группировку sui generis. Это убеждение еще более крепнет, когда замечаешь, что здесь не исключительный и изолированный случай, а что подобные единства складывались в ходе истории жизни неоднократно. Приведем только два примера, опять-таки не выходя за пределы млекопитающих.
В третичный период, учит геология, часть плацентарного биота, находившегося тогда в состоянии глубокой эволюции, оказалась отрезанной морем в южной половине Американского континента. Как же реагировал этот черенок на свою изоляцию? Точно так же, как реагирует черенок растения, то есть воспроизводя в меньшем масштабе рисунок ствола, от которого был отделен. Он начал выпускать отростки своих псевдохоботных, своих псевдогрызунов, своих псевдолошадей, своих псевдообезьян (широконосые обезьяны)... Целый биот в уменьшенном виде (подбиот) внутри первоначального биота!
Теперь второй пример, касающийся сумчатых.
Судя по их сравнительно примитивной форме размножения, а также по их современному географическому распространению, явно несплошному и остаточному, сумчатые (аплацентарные) представляют собой особый ярус у основания млекопитающих. Они, очевидно, достигли расцвета раньше плацентарных и образовали раньше их свой собственный биот. В целом, за исключением нескольких странных типов (вроде псевдо-«Machairodus», ископаемые останки которого недавно найдены в Патагонии)5, этот биот сумчатых бесследно исчез. Но зато один из его подбиотов, развившийся еще до третичного периода и случайно сохранившийся опять же вследствие изоляции в Австралии, до сих пор вызывает восхищение натуралистов четкостью своих контуров и своим совершенством. Как всем известно, Австралия при открытии ее европейцами была населена лишь сумчатыми6, но зато сумчатыми всех размеров, всех зон обитания, всех форм - сумчатыми травоядными и бегающими, хищными сумчатыми, насекомоядными сумчатыми, сумчатыми крысами, сумчатыми кротами и т. д. Нельзя себе представить более яркий пример способности, присущей каждой филе, к дифференциации в своего рода замкнутый, физиологически полный организм.
Установив это, подымимся выше, чтобы рассмотреть обширную систему двух биотов - плацентарного и аплацентарного, вместе взятых. Зоологи очень быстро заметили, что у всех форм, входящих в эти две группы, коренные зубы состоят в основном из трех бугорков, зацепляющихся от одной челюсти к другой, сверху вниз. Черта, незначительная сама по себе, но тем более интригующая своим постоянством.
Как объяснить универсальность столь случайной черты? Ключ к отгадке был дан открытием, сделанным в некоторых юрских отложениях в Англии. В средней юре на какое-то мгновение нам приоткрывается первая пульсация млекопитающих - мир маленьких зверьков, не больше крыс или землероек. И вот у этих-то мелких, но уже чрезвычайно разнообразных животных тип зубов еще не закреплен, как в нынешней природе. Среди них уже можно различить и трехбугорчатый тип. Но наряду с ним в развитии и способах противоположения бугорков на коренных зубах можно наблюдать и всякие другие комбинации. И эти другие комбинации уже давно устранены! Напрашивается вывод. За исключением, может быть, утконоса и ехидны (этих яйценосных парадоксальных форм, в которых предполагали продолжение многобугорчатых), все ныне живущие млекопитающие ведут начало от одного узкого пучка. Все вместе они представляют собой (в развитом состоянии) лишь один из многочисленных побегов, на которые делится юрская мутовка млекопитающих - побег трехбугорчатых7...
Здесь мы почти достигли предела того, что можно видеть сквозь толщу прошлого. Ниже, кроме вероятного существования в самом конце триаса еще одной мутовки, к которой относятся многобугорчатые, история млекопитающих теряется во мраке.
По крайней мере около этого времени и выше их группа, естественно изолированная из-за отрыва черешка, выделяется с достаточной четкостью и индивидуальностью, чтобы мы могли ее взять в качестве практической единицы «эволюционной массы».
Назовем эту единицу пластом.
Нам тут же придется ею пользоваться.
б) Пласт пластов - четвероногие
Когда необходимо измерить расстояние до туманностей, астрономы пользуются световыми годами. В свою очередь, если мы хотим, отправляясь от млекопитающих, расширить и продолжить вниз наше видение древа жизни, нам надо отсчитывать пластами.
Для начала возьмем пласты пресмыкающихся вторичного периода.
Когда ниже юрского периода мы теряем из виду ветвь млекопитающих, то она не рассеивается в какой-то пустоте. Ее покрывает и обволакивает густая листва жизни совершенно иного вида: динозавры, птерозавры, ихтиозавры, крокодилы и масса других чудищ, менее знакомых для людей, не сведущих в палеонтологии. В этом ансамбле зоологические расстояния между формами явно более велики, чем между отрядами млекопитающих. Однако три особенности бросаются в глаза. Прежде всего здесь мы имеем дело с разветвленной системой. В этой системе, далее, ветви выступают на уже зрелой или даже конечной стадии распускания. Наконец, вся группа в целом представляет собой не что иное, как огромный и, может быть, сложный биот. Тут - травоядные, зачастую гигантских размеров. Там - их спутники и тираны - плотоядные, массивные или прыгающие. Здесь - летающие, с перепонками летучих мышей или перьями птиц. И, наконец, плавающие, такие же обтекаемые, как и дельфины.
На расстоянии этот мир пресмыкающихся нам представляется более сжатым, чем мир млекопитающих, но его длительность, измеряемая конечной экспансией и усложнением, может считаться по крайней мере равной. Во всяком случае, он также исчезает в глубинах времен. К середине триаса динозавров еще можно заметить. Но они едва выступают из другого пласта, почти пришедшего к своему закату, - пласта пермских пресмыкающихся, для которых особенно характерны зверообразные ящеры.
Толстые и бесформенные, к тому же редкие в наших музеях, зверообразные значительно менее популярны, чем диплодоки и игуанодоны. Но это не мешает им приобретать все большее значение в зоологии. Если раньше на них смотрели как на странные, диковинные существа, узко ограниченные пределами Южной Африки, то теперь окончательно признано, что они одни представляют целую стадию жизни континентальных позвоночных. В определенный период, до динозавров, до млекопитающих, именно они занимали всю не покрытую морем территорию и владели ею. Или вернее будет сказать, что эти ящеры, крепко стоявшие на своих хорошо сочлененных конечностях, зачастую обладавшие «корневидными» зубами, были первыми четвероногими, прочно утвердившимися на суше.
В тот момент, когда мы их обнаруживаем, они уже изобилуют в самых причудливых формах - с рогами, с гребнями, с бивнями, что (как всегда!) указывает на группу, достигшую конца своей эволюции. Фактически же при всей своей внешней причудливости это довольно монотонная группа, в которой еще не различаются ясно прожилки настоящего биота. И все же эта группа поражает развертыванием вширь и потенциальными возможностями своей мутовки. С одной стороны, неподвижные черепахи. С другой - чрезвычайно прогрессивные по своей подвижности и строению черепа типы, из которых, как мы имеем все основания предполагать, и выступил давно дремавший стебель млекопитающих.
А дальше новый «провал». На этих расстояниях, под тяжестью прошлого отрезки длительности быстро сжимаются. Когда у основания пермского периода и ниже его мы снова различаем поверхность обитаемой Земли, то она оказывается заселенной лишь земноводными, ползающими по тине. Земноводные - изобилие приземистых или змеевидных тел, среди которых зачастую трудно отличить личинки от взрослых особей; голая или покрытая чешуей кожа; трубчатые или в виде мозаики косточек позвонки... И здесь, согласно общему правилу, нам удается схватить уже только высокодифференцированный мир, почти пришедший к своему концу. Сколько еще пластов мы, может быть, смешиваем в этом кишении, исследуя осадки, толщину и громадность истории которых мы еще плохо осознаем. Но по крайней мере достоверно одно - на этой стадии мы схватываем животную группу, которая начинает выходить из воды-кормилицы, в которой она образовалась.
Но уже в этом самом начале своей субвоздушной жизни позвоночные демонстрируют перед нами одну удивительную особенность, над которой следует поразмыслить. У всех у них «одна и та же структура скелета; в особенности она тождественна (не говоря уже о поразительных соответствиях черепа) в отношении числа и плана двигательных конечностей. В чем причина этой схожести?
То, что все земноводные, пресмыкающиеся и млекопитающие имеют четыре, и только четыре, лапы, может в крайнем случае объясняться просто стремлением к наиболее простой форме передвижения (однако насекомые никогда не имеют менее шести ног...). Но как объяснить чисто механическими причинами совершенно одинаковую структуру этих четырех отростков? Впереди - единственная плечевая кость, за ней две кости предплечья, затем пять лучей руки?.. Не представляет ли это еще раз одну из тех случайных комбинаций, которые были открыты и реализованы лишь однажды? Здесь снова поневоле напрашивается вывод, уже сделанный в случае трехбугорчатости млекопитающих. Несмотря на свое чрезвычайное разнообразие, обладающие легкими наземные животные представляют собой не что иное, как вариации, нагроможденные на совершенно частном решении жизни.
Таким образом, огромный и сложный веер ходячих позвоночных, будучи продолжен к своим истокам, снова свертывается и смыкается в один луч.
Один-единственный черешок замыкает и определяет у его основания пласт пластов - мир четвероногих.
в) Ветвь позвоночных
В случае млекопитающих мы сумели схватить мутовку, из которой выделился и устремился вперед побег «трехбугорчатых». Что же касается происхождения земноводных, то тут наука не достигла таких успехов. Однако можно без колебаний указать ту единственную область жизни, в которой среди других испробованных комбинаций могли образоваться четвероногие. Они должны были возникнуть где-то среди рыб с дольчатыми и «членообразными» плавниками, некогда жизнеустойчивый пласт которых ныне существует лишь в виде нескольких живых ископаемых: двоякодышащих (или рыб, обладающих легкими) и - совсем свежий сюрприз - кистеперой рыбы (Crossopterigien), недавно выловленной в южных морях.
Поверхностно «гомогенизированные» путем механической адаптации к плаванию, рыбы (лучше сказать рыбообразные) невероятно сложны в своей совокупности. Сколько пластов, особенно здесь, соединено и смешано под одним и тем же названием?.. Сравнительно молодых пластов, развившихся в океане в ту же эпоху, когда на континентах распространились пласты четвероногих. И еще более многочисленных старых пластов, заканчивающихся очень низко, у силура, основной мутовкой, от которой, на наш взгляд, расходятся два главных побега: бесчелюстные рыбообразные, с единственной ноздрей, представленные в нынешней природе одной лишь миногой; и челюстные рыбообразные, с двумя ноздрями, от которых пошло все остальное.
После всего вышесказанного о последовательном развитии земных форм я не буду пытаться затрагивать и расчленять этот другой мир. Я постараюсь лучше обратить внимание на факт другого порядка, с которым мы встречаемся здесь впервые. Самые древние рыбы, которых мы знаем, в большинстве случаев облечены в мощный, даже чересчур мощный панцирь8. Но под этой первой, очевидно довольно безуспешной, попыткой упрочения извне скрывается скелет еще целиком из хрящей. По мере следования за ними вниз позвоночные выступают перед нами все менее окостенелыми внутри. И этим объясняется тот факт, что даже в неповрежденных отложениях мы полностью теряем их следы. Но в данном частном случае проявляется общее явление, имеющее важнейшее значение. Какую бы живую группу мы ни рассматривали, в глубине она всегда тонет в сфере мягкого. Безошибочный способ скрыть свой черешок...
Ниже девонского периода рыбообразные вступают в своего рода фетальную или личиночную фазу, не поддающуюся окаменению. Не доживи случайно до нас странный ланцетник, мы бы не имели никакого понятия о многочисленных этажах, из которых, по-видимому, строился тип хордовых, до тех пор пока он стал в состоянии заполнить воду, а в последующем и сушу.
Так заканчивается и ограничивается у основания большой пустотой громадное сооружение всех четвероногих и всех рыб - ветвь позвоночных.
г) Остальная жизнь
Ветвь - это наиболее широкий тип оформленной совокупности, еще признаваемый систематикой внутри биосферы. Помимо позвоночных, основную крону жизни образуют еще только две ветви - ветвь червей и членистоногих и ветвь растений. Приобретая прочность - одна с помощью хитина и известняка, другая с помощью целлюлозы, - они также сумели вырваться из плена воды и широко распространиться в воздушной среде. В результате растения и насекомые в нынешней природе перемешались и борются с костистыми животными за то, кто больше займет места в мире.
Для каждой из этих двух ветвей можно было бы повторить анализ (но я не стану этого делать), предпринятый в предыдущих параграфах о позвоночных. Вверху - молодые группы, богатые легкими мутовками. Глубже - пласты с более четко выраженными, но менее густыми разветвлениями. В самом низу - погружение в мир химически неустойчивых форм. Та же самая общая картина развития. Но, так как в данном случае ветви, конечно, старше, то здесь и большее усложнение; а в случае насекомых - крайние формы социализации.
Не может быть сомнения, что в бездне времен эти различные линии сходятся к какому-то общему полюсу рассеивания. Но задолго до того, как смыкаются хордовые, кольчатые черви и растения (две первые ветви, очевидно, среди многоклеточных; эти последние и растения - только на уровне одноклеточных существ), их стволы исчезают в скоплении причудливых форм - губок, иглокожих, полипов... Все это - наброски ответов на проблему жизни. Пучок неудавшихся ветвей.
Все они, несомненно, возникли (хотя мы и не можем сказать, каким образом, - столь глубоким из-за действия длительности стал разрыв) из другого, невероятно древнего и многообразного мира - инфузорий, различных простейших, бактерий, свободных клеток, обнаженных или покрытых щитком, у которых царства жизни смешались, и систематика бессильна. Животные это или растения? Эти слова более не имеют смысла. Наслаивание пластов и ветвей, или «мицелий», перепутанных волокон, как у гриба? Этого мы уже не знаем. Не знаем мы также, из чего это все зародилось. Начиная с докембрия и одноклеточные в свою очередь теряют свой остов из кремния или известняка. И тем самым в мягкости тканей и в метаморфозе первородного ила окончательно исчезают с наших глаз корни древа жизни.
Б. Размеры
Вот и набросана, хотя и весьма схематично, структурная картина форм, собранных и классифицированных упорным трудом натуралистов, начиная с Аристотеля и Линнея. В ходе описания мы уже старались дать почувствовать огромную сложность этого возрожденного нами мира. Перед лицом всей совокупности нам надо, однако, последним усилием видения более ясно осознать эту громадность размеров.
Сам по себе наш рассудок постоянно склонен не только прояснять (что составляет его функцию), но и сужать и укорачивать реальности, к которым прикасается. Устав, он сгибается под тяжестью расстояний и множеств. Так что, обрисовав с грехом пополам экспансию жизни, нам теперь надо представить себе элементы нашей схемы в их истинных размерах - как по численности, так и по объему и по длительности.
Попытаемся это сделать.
Прежде всего по численности. Для простоты наш эскиз живого мира делался с помощью широких коллективных групп - семейств, отрядов, биотов, пластов, ветвей... Но. пользуясь этими различными единицами, думали ли мы о множествах, с которыми реально имели дело? Пусть тот, кто хочет осмыслить или описать эволюцию, прежде всего отправится побродить в один из тех четырех-пяти крупнейших музеев мира, в которых (ценой усилий, героизм и духовное значение которых будут однажды оценены) легион путешественников сумел в сжатом виде в нескольких залах представить весь спектр жизни. И пусть он там, не обращая внимания на названия, как следует проникнется тем, что его окружает. Тут - мир насекомых, где полноценные виды исчисляются десятками тысяч. Там - моллюски, другие тысячи, бесконечно разнообразные по рисунку прожилок и форме завитков. Затем идут рыбы, с такими же неожиданными, капризными и пестрыми формами, как у бабочек. Далее птицы, едва ли менее фантастические, самого разного профиля, со всякими клювами, всех расцветок. Потом антилопы, всех мастей, с разными осанками, со всевозможными диадемами и т. д. и т. д. Какое многообразие, какой, порыв, какое бурление под каждым из этих названий, которые вызывали в нашем воображении какую-нибудь дюжину весьма привычных форм! А ведь перед нами здесь только ныне живущие. Что, если бы мы могли видеть всех остальных... Во все эпохи Земли, на всех стадиях эволюции другие музеи зарегистрировали бы такое же бурление, такое же буйное изобилие. Все эти сотни тысяч названий, внесенных в каталоги нашей систематики, не представляют и миллионной доли листьев, произраставших до сего дня на древе жизни.
Теперь по объему. То есть каково количественное соотношение между различными зоологическими и ботаническими группами в природе? Какова материально доля каждой из них в общей совокупности организованных существ?
Чтобы дать общее представление об этой пропорции, я воспроизвожу здесь (рис. 2) выразительный рисунок, которым один ученый натуралист г. Кено представил, по самым последним данным науки, карту животного царства с его основными подразделениями. Эта карта больше позиционная, чем структурная, но она точно отвечает на поставленный вопрос.

Рис. 2. Древо жизни, по Кено (изд. Массон и К°). На этом символическом рисунке каждая главная долька (или гроздь) соответствует одному пласту, по крайней мере столь же важному (морфологически и количественно), как пласт, составляемый всеми млекопитающими. Ниже линии АВ - формы, живущие в воде; выше ее - живущие в воздушной среде.
Рассмотрим эту схемуiv. Не правда ли, при первом же взгляде наш рассудок испытывает шок, подобно тому шоку, который мы чувствуем, когда астроном показывает Солнечную систему как простую звезду, все наши звезды - как один Млечный Путь, а наш Млечный Путь - как атом среди других Галактик?.. Млекопитающие, в которых обычно воплощается для нас идея и образ «животного», жалкая долька, поздно возникшая на стволе жизни! А зато что делается вокруг них! А рядом? А ниже?.. Какое обилие соперничающих типов, о существовании, величине, множестве которых мы и не подозревали! Таинственные существа, которых мы могли случайно видеть скачущими среди сухих листьев или ползающими по песку, ни разу не задаваясь вопросом, что они означают и откуда взялись. Существа, незначительные по своим размерам и ныне, быть может, по числу... Эти оставленные в пренебрежении формы теперь предстают в своем истинном свете. По богатству своих разновидностей, по времени, которое потребовалось природе для их порождения, каждая из них представляет собой столь же важный мир, как и наш. Количественно (я подчеркиваю это) мы - лишь одни из них и притом возникшие последними.
Наконец, по длительности. Здесь, как обычно, нашему воображению трудно восстановить подлинную картину. Как я уже отмечал, еще более неумолимо, чем горизонты пространства, в нашей перспективе сближаются и налезают друг на друга плоскости прошлого. Как же их разделить?
Чтобы придать глубинам жизни их истинный рельеф, полезно для начала вернуться к тому, что я выше назвал пластом млекопитающих. Поскольку этот пласт относительно молод, у нас есть некоторое представление о времени, потребовавшемся для его развития начиная с момента, когда он явственно поднялся над пресмыкающимися в конце мелового периода. Весь третичный период и еще немного. Каких-нибудь 80 миллионов лет. Допустим теперь, что на оси одной и той же зоологической ветви пласты образуются периодически, как сучки вдоль ствола хвойного дерева; так что максимумы их расцвета (единственно четко регистрируемые) в случае позвоночных следуют друг за другом на расстоянии в 80 миллионов лет. Чтобы получить величину длительности одного зоологического интервала, достаточно помножить 80 миллионов лет на число пластов в данном интервале. Так, между млекопитающими и началом четвероногих имеется не менее трех пластов. Числа становятся внушительными. Но они достаточно хорошо согласуются с представлениями, которые вырабатывает геология, о громадности триасового, пермского и каменноугольного периодов.
Можно применить и другой, более приблизительный метод, основанный на сравнении ветвей. Внутри одного пласта (возьмем снова пласт млекопитающих) мы в состоянии примерно определить среднее отклонение форм между собой - для всего этого рассеивания потребовалось, повторяем, около 80 миллионов лет. После этого сравним между собой млекопитающих, насекомых и высшие растения. Если три ветви, на конце которых цветут эти три группы, расходятся из одного и того же корня, а не возникли раздельно на одном «мицелии» (что возможно), то какое же требовалось время, сколько периодов, чтобы создать эти гигантские разрывы между типами! Здесь уже зоология бросает вызов данным, приводимым геологией. Прошло всего лишь 1 500 миллионов лет со времени самых древних следов углерода в осадках, решили физики, измерив процент содержания свинца в одном радиоактивном минерале докембрия. Но не предшествуют ли первые организмы этим первым следам? И потом в случае разногласия при подсчете возраста Земли - на какой из двух хронометров следует положиться? На медленность разложения радия? Или на медленность «сложения» живой материи?
Если простой секвойе для своего полного роста требуется пять тысяч лет (и еще никто не видел, чтобы секвойя погибла своей естественной смертью), то каков же должен быть полный возраст древа жизни.
В. Очевидность
И вот теперь это дерево перед нами. Странное это дерево. Можно сказать, негатив дерева, ибо в отличие от гигантов наших лесов его ветви, его ствол обнаруживают себя перед нами лишь пустотами все большего диаметра. С виду - застывшее дерево, столь длительным кажется нам раскрытие почек, которые мы всегда будем узнавать лишь полураскрытыми. Но тем не менее дерево, ясно очерченное своей многоярусной листвой наблюдаемых видов. В своих основных чертах и размерах оно стоит перед нами, покрывая Землю. Рассмотрим его получше, прежде чем проникать в секрет его жизни. Ибо уже из простого созерцания его внешних форм можно извлечь один важный урок - чувство его очевидности.
В мире еще встречаются люди, подозрительно и скептически настроенные в отношении эволюции. Зная природу и натуралистов лишь по книгам, они полагают, что борьба вокруг трансформизма все еще продолжается, как во времена Дарвина. А поскольку в биологии идут дискуссии относительно механизма видообразования, они воображают, что эта наука сомневается или еще могла бы сомневаться, не отрицая самой себя, насчет факта и реальности такого развития.
Но положение совершенно иное.
Может показаться удивительным, что в этой главе, посвященной последовательной цепи развития организованного мира, я ни разу не упомянул о продолжающихся оживленных спорах по вопросу о различии между сомой и зародышевым веществом, о существовании и функциях генов, о передаваемости или непередаваемости приобретенных признаков. Но это потому, что на данном этапе моего исследования эти вопросы меня непосредственно не интересуют.
Для подготовки естественных рамок для антропогенеза и колыбели для человека - я хочу сказать, для гарантирования существенной объективности эволюции - необходимо и достаточно одно, чтобы общий филогенез жизни (каковы бы ни были его процесс и движущая пружина) был для нас столь же отчетливо различим, как и индивидуальный ортогенез, через который на наших глазах проходит каждое живое существо, чему мы нисколько не удивляемся.
Так вот, доказательство этого общего развития биосферы образуется в нашем уме почти механически и неотвратимо тем материальным рисунком, к которому мы неизбежно приходим при каждой новой попытке установить, штрих за штрихом, контуры и структуру организованного мира.
Никому не придет в голову подвергнуть сомнению вихревое происхождение спиральных туманностей; или последовательное приращение частиц у кристалла или сталагмита; или наращивание пучков волокон вокруг оси стебля. Некоторые геометрические построения, на наш взгляд совершенно неподвижные, являются следом и верным признаком кинематики. Как же можно хоть на миг усомниться в эволюционном происхождении живого слоя Земли?
Уступая силе анализа, жизнь расслаивается. Она до бесконечности расчленяется в анатомически и физиологически связную систему вставленных друг в друга вееров9. Едва наметившиеся микровеера подвидов и рас. Уже более широкие веера видов и родов. Все более и более обширные веера биотов, затем пластов и далее ветвей. И, наконец, вся совокупность животного и растительного царств, образующая путем ассоциации один гигантский биот, быть может, берущий начало как один простой побег от мутовки, находящейся в глуби мегамолекулярного мира. Жизнь - простая ветвь чего-то другого...
Сверху донизу, от самого большого до самого малого - одна и та же видимая структура, рисунок которой, подкрепленный самим распределением теней и пустот, принимает все более четкий характер и продолжается (независимо от всякой гипотезы!) в почти самопроизвольном расположении непредвиденных элементов, добавляющихся ежедневно. Каждая вновь открытая форма находит свое естественное место, фактически в данных рамках ни одна из них не является абсолютно «новой». Что еще нужно для убеждения, что все это родилось, что все это выросло?..
После этого мы можем еще годами спорить о том, как возник этот огромный организм. Убеждаясь все больше в ошеломляющей сложности его строения, мы испытываем головокружение. Как совместить этот постоянный рост с детерминизмом молекул, со слепой игрой хромосом, с кажущейся неспособностью индивидуальных приобретений передаваться по наследству? Иначе говоря, как совместить внешнюю, «финалистскую» эволюцию фенотипов с внутренней, механистической эволюцией генотипов?.. Разбирая машину на части, мы перестаем понимать, как она может двигаться вперед. Может быть и так. Но тем не менее машина перед нами, и она движется. Если химия еще не может сказать ничего вразумительного о способе образования гранитов, то можно ли сомневаться в том, что континенты беспрестанно становятся все более гранитными?
Как все в универсуме, где время окончательно (я к этому еще вернусь) стало четвертым измерением, жизнь не может не быть величиной эволюционной по своей природе или размерам. Физически и исторически она соответствует некоторой функции X, определяющей положение каждого живого существа в пространстве, в длительности и по форме. Вот фундаментальный факт, который требует объяснения, но очевидность которого уже не нуждается в проверке и гарантирована от опровержения опытом в последующем.
На этой ступени обобщения можно сказать, что «трансформистский вопрос» более не существует, он окончательно решен. Отныне, чтобы поколебать наше убеждение в реальности биогенеза, надо было бы подрывая всю структуру мира, вырвать с корнем древо жизни10.
1 Под предлогом, что термин «ортогенез» употреблялся в различных спорных или ограниченных значениях, или потому, что он имеет метафизический привкус, некоторые биологи хотели бы его попросту устранить. Я же, напротив, твердо убежден, что это слово имеет существенное значение и незаменимо, когда надо отметить и подчеркнуть очевидное свойство живой материи образовывать систему. «внутри которой ее члены, как показывает опыт. следуют друг за другом по все возрастающим значениям центросложности».
2 Если бы наши аппараты (автомобили, самолеты и т. д.) путем какого-либо катаклизма оказались под землей и превратились в «ископаемые», то будущие геологи, открыв их, получили бы такое же впечатление, как мы от птеродактиля: представленные единственно своими последними марками, эти продукты нашей изобретательности показались бы им созданными без фазы пробных поисков - сразу готовыми и законченными.
3 Как будет замечено далее в связи с «моногенизмом», мы не случайно не можем выйти за определенный предел точности («разделения») в нашем восприятии очень далекого прошлого (каждый раз по случайным причинам - см. Курно...). Во всех направлениях (в направлении очень древнего и очень малого, но также в направлении очень большого и очень медленного) наша зоркость слабеет; за пределами некоторого радиуса мы более ничего не различаем.
4 Так называют в противоположность аплацентарным (или сумчатым) млекопитающих, у которых зародыш благодаря наличию специальной оболочки - плаценты, которая его защищает и питает, может до полного созревания оставаться в утробе матери.
5 Махайрод, или саблезубый тигр. Эта крупная кошка, весьма распространенная в конце третичного и начале четвертичного периодов, имеет странный дубликат в лице жившего в плиоцене хищного сумчатого Южной Америки.
6 Кроме одной группы грызунов и прибывших последними человека и его собаки.
7 которых можно также назвать «семипозвонковыми», потому что по столь же неожиданному и столь же замечательному совпадению все они имеют семь шейных позвонков, какова бы ни была длина их шеи.
8 Без этого окостеневшего покрова от них ничего бы и не осталось, и мы бы о них ничего не знали.
9 Разумеется, в этой игре вееров стыки можно было бы наметить иначе, чем это сделано мною, - в частности отведя больше места параллелизму и схождению. Например, четвероногие могут рассматриваться как пучок, состоящий из ряда побегов, вышедших из различных мутовок, но одинаково пришедших к формуле четвероножия. Эта полифилетическая схема, на мой взгляд, менее верно выражает факты. Но она нисколько не меняет моего основного положения, а именно, что жизнь выступает как органически сочлененный ансамбль, с очевидностью выражающий феномен роста.
10 Действительно, постольку поскольку эволюционизм выражает простую невозможность для нас воспринимать в опыте любое существо (живое или неживое) иначе, как в пространственно-временном ряду, он давно перестал быть гипотезой и стал условием (мерности), которому отныне должны отвечать в физике и биологии все гипотезы. В настоящее время биологи и палеонтологи еще спорят о конкретных путях и особенно о механизме превращений жизни: имеет ли место при возникновении новых признаков господство случая (неодарвинизм) или же факт изобретения (неоламаркизм). Но с тем общим и основным фактом, что органическая эволюция происходит как в случае любого живого существа в отдельности, так и в случае жизни в целом, ныне согласны все ученые; и это по той простой причине, что иначе они вообще не могли бы заниматься наукой... Можно лишь сожалеть (и удивляться), что. несмотря на ясность фактов. единодушие еще не доходит до признания того, что «галактика» жизненных форм вырисовывает (как это допущено нами) широкое «ортогенетическое» движение свертывания в направлении все большего возрастания сложности и сознания (см. Заключение в конце книги).
i Имеется в виду «парадокс трансформизма», раскрытый Тейяром еще за несколько лет до того. как он начал писать «феномен человека». Начало, «черешок» каждой филы или эволюционного ствола само еще не является ее типичным компонентом и обычно сливается в восприятии палеонтолога или эволюциониста с филой-источником. либо же вообще ускользает от внимания исследователя в силу скудости сохраняющегося и дошедшего до нас материала (Р. Teilhard de Chardin. Le paradoxe transformiste. «Rev. Quest. Sci.». 1925, № 7. p. 53-80).
ii Здесь Тейяр ссылается на идею французского философа-феноменолога. математика и экономиста Ангуана Огюстена Курно (1801-1877). изложенную в его «Очерке об основах наших знаний и об особенностях философской критики» («Essai sur les fondements de nos connaissance et sur les caracteres de la critique philosophique», Paris. 1851).
iii Имеется в виду эволюционно-палеонтологическое направление, созданное в США Генри Фейрфилдом Осборном (1857-1953), автором многих работ по филогенезу млекопитающих. Цитируемая в тексте идея высказывалась Осборном в 10-30-х годах XX в., когда он отошел от первоначально отстаивавшегося им ламаркизма и предпринял попытку синтезировать (для объяснения эволюции) виталистические и энергистские концепции с элементами неодарвинизма. Осборновская концепция «аристогенов» или «генов прогресса» послужила одним из источников учения Тейяра о радиальной энергии.
iv В своей общей схеме эволюции Тейяр во многом опирается на работы одного из первых французских генетиков Люсьена Кено (1866-1951). развивавшего концепцию преадаптации и «антислучайности» в противовес якобы основанному лишь на факторе «случайности» дарвиновскому естественному отбору. Приводимая Тейяром схема Кено была в 30-40-е годы принята практически всеми французскими эволюционистами как классическая и имеет то преимущество перед многими другими «филогенетическими деревьями», что в ней иллюстрирована множественность направлений освоения суши и воздушной среды в различных эволюционных стволах животного мира. В § 2 «Подъем сознания» главы III («Мать-Земля») данного раздела Тейяр применяет концепцию «антислучайности» Кено для разделения двух «зон эволюции - дарвинистской. где закономерность пробивается сквозь массу случайностей, и ламаркистской. где она «ощутимо доминирует». К второй зоне Тейяр относит прежде всего эволюцию человека: практически различие между «зонами» соответствует различию между биологической и культурной эволюцией. Тейяр вслед за Кено явно недооценивает закономерного характера эволюции под действием естественного отбора.